| Size | Price | Stock | Qty |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| Other Sizes |
|
































Purity: ≥98%
AR-C155858 is a novel, selective and potent inhibitor of monocarboxylate transporters MCT1 and MCT2 with Kis of 2.3 nM and 10 nM, respectively. It binds to an intracellular site involving transmembrane helices 7-10. Inhibitor titrations of L-lactate transport by MCT1 in rat erythrocytes were used to determine the Ki and number of AR-C155858-binding sites (Et) on MCT1 and the turnover number of the transporter (kcat). Derived values were 2.3+/-1.4 nM, 1.29+/-0.09 nmol per ml of packed cells and 12.2+/-1.1 s-1 respectively.
| Targets |
MCT1/2 (monocarboxylate transporter) (Ki = 2.3 nM and 10 nM)
|
|---|---|
| ln Vitro |
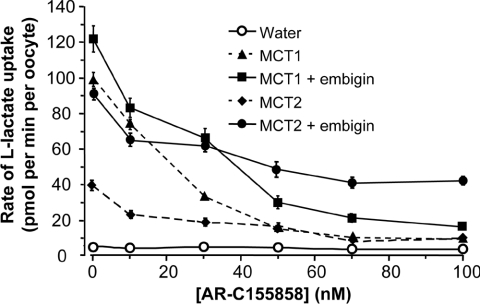
The MCT1/MCT2 C-terminal chimera is inhibited by AR-C155858 (10 nM–100 nM) [1]. MCT2 is inhibited by AR-C155858, with a 70% inhibition at 10 nM and a progressive increase in inhibition that is only consistent with Ki values much below 10 nM. In Xenopus laevis oocytes, AR-C155858 suppresses MCT1 expression in a concentration- and time-dependent manner[2].
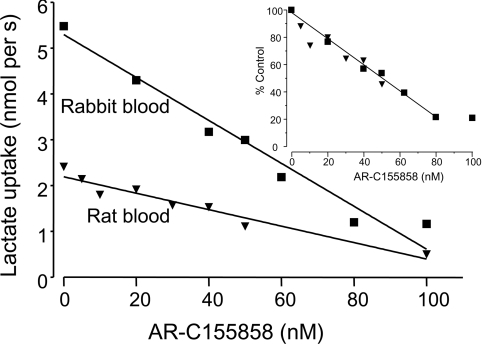
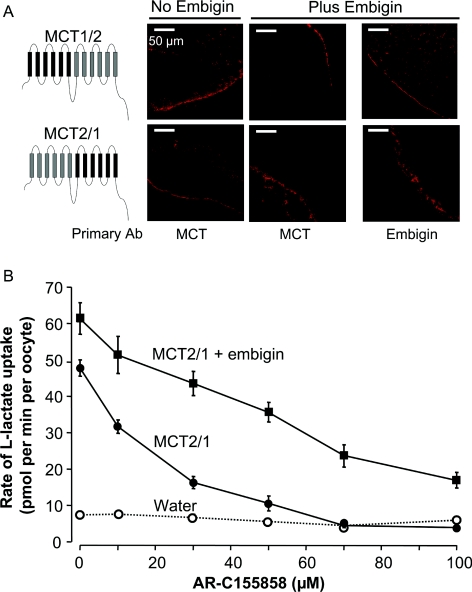
In mammalian cells, MCTs (monocarboxylate transporters) require association with an ancillary protein to enable plasma membrane expression of the active transporter. Basigin is the preferred binding partner for MCT1, MCT3 and MCT4, and embigin for MCT2. In rat and rabbit erythrocytes, MCT1 is associated with embigin and basigin respectively, but its sensitivity to inhibition by AR-C155858 was found to be identical. Using RT (reverse transcription)–PCR, we have shown that Xenopus laevis oocytes contain endogenous basigin, but not embigin. Co-expression of exogenous embigin was without effect on either the expression of MCT1 or its inhibition by AR-C155858. In contrast, expression of active MCT2 at the plasma membrane of oocytes was significantly enhanced by co-expression of exogenous embigin. This additional transport activity was insensitive to inhibition by AR-C155858 unlike that by MCT2 expressed with endogenous basigin that was potently inhibited by AR-C155858. Chimaeras and C-terminal truncations of MCT1 and MCT2 were also expressed in oocytes in the presence and absence of exogenous embigin. L-Lactate Km values for these constructs were determined and revealed that the TM (transmembrane) domains of an MCT, most probably TM7–TM12, but not the C-terminus, are the major determinants of L-lactate affinity, whereas the associated ancillary protein has little or no effect. Inhibitor titrations of lactate transport by these constructs indicated that embigin modulates MCT2 sensitivity to AR-C155858 through interactions with both the intracellular C-terminus and TMs 3 and 6 of MCT2. The C-terminus of MCT2 was found to be essential for its expression with endogenous basigin. [1] In the present study we characterize the properties of the potent MCT1 (monocarboxylate transporter 1) inhibitor AR-C155858. Inhibitor titrations of L-lactate transport by MCT1 in rat erythrocytes were used to determine the Ki value and number of AR-C155858-binding sites (Et) on MCT1 and the turnover number of the transporter (kcat). Derived values were 2.3+/-1.4 nM, 1.29+/-0.09 nmol per ml of packed cells and 12.2+/-1.1 s-1 respectively. When expressed in Xenopus laevis oocytes, MCT1 and MCT2 were potently inhibited by AR-C155858, whereas MCT4 was not. Inhibition of MCT1 was shown to be time-dependent, and the compound was also active when microinjected, suggesting that AR-C155858 probably enters the cell before binding to an intracellular site on MCT1. Measurement of the inhibitor sensitivity of several chimaeric transporters combining different domains of MCT1 and MCT4 revealed that the binding site for AR-C155858 is contained within the C-terminal half of MCT1, and involves TM (transmembrane) domains 7-10. This is consistent with previous data identifying Phe360 (in TM10) and Asp302 plus Arg306 (TM8) as key residues in substrate binding and translocation by MCT1. Measurement of the Km values of the chimaeras for L-lactate and pyruvate demonstrate that both the C- and N-terminal halves of the molecule influence transport kinetics consistent with our proposed molecular model of MCT1 and its translocation mechanism that requires Lys38 in TM1 in addition to Asp302 and Arg306 in TM8 [Wilson, Meredith, Bunnun, Sessions and Halestrap (2009) J. Biol. Chem. 284, 20011-20021] [2]. |
| ln Vivo |
Purpose: Monocarboxylate transporter (MCT) inhibition represents a potential treatment strategy for γ-hydroxybutyric acid (GHB) overdose by blocking its renal reabsorption in the kidney. This study further evaluated the effects of a novel, highly potent MCT inhibitor, AR-C155858, on GHB toxicokinetics/toxicodynamics (TK/TD). [2]
Methods: Rats were administered GHB (200, 600 or 1500 mg/kg i.v. or 1500 mg/kg po) with and without AR-C155858. Breathing frequency was continuously monitored using whole-body plethysmography. Plasma and urine samples were collected up to 8 h. The effect of AR-C155858 on GHB brain/plasma partitioning was also assessed. [2] Results: AR-C155858 treatment significantly increased GHB renal and total clearance after intravenous GHB administration at all the GHB doses used in this study. GHB-induced respiratory depression was significantly improved by AR-C155858 as demonstrated by an improvement in the respiratory rate. AR-C155858 treatment also resulted in a significant reduction in brain/plasma partitioning of GHB (0.1 ± 0.03) when compared to GHB alone (0.25 ± 0.02). GHB CLR and CLoral (CL/F) following oral administration were also significantly increased following AR-C155858 treatment (from 1.82 ± 0.63 to 5.74 ± 0.86 and 6.52 ± 0.88 to 10.2 ± 0.75 ml/min/kg, respectively). [2] Conclusion: The novel and highly potent MCT inhibitor represents a potential treatment option for GHB overdose. [2] |
| Enzyme Assay |
Measurement of MCT1 activity in rat erythrocytes [2]
L-Lactate transport into rat erythrocytes was measured by monitoring the change in extracellular pH with a pH-sensitive electrode as described previously. The cells were used at 3.5% or 7% haematocrit in lightly buffered saline medium supplemented with 5 μM DIDS and 100 μM acetazolamide to prevent bicarbonate/CO2-mediated proton movements. The erythrocytes were pre-incubated for 1 h at room temperature (22–25 °C) with or without AR-C155858 at the required concentration prior to assaying lactate transport. This was performed at 6 °C with substrate uptake initiated by addition of 10 mM L-lactate. Initial rates of transport were calculated by first-order regression analysis of the time course of pH change and converted into nmol of H+ per min by determining the pH change induced by small additions of standardized NaOH. Measurement of MCT transport activity in Xenopus oocytes [2] cRNA was prepared by in vitro transcription from the appropriate linearized pGHJ plasmid and injected into X. laevis oocytes as described previously [33]. For most assays 20 ng of cRNA was injected, but for [14C] kinetic assays, injection quantity was adjusted to ensure that uptake was linear with time. Further details can be found in Supplementary Table S2 (at http://www.BiochemJ.org/bj/425/bj4250523add.htm). Controls received the equivalent volume (9.2 nl) of water. Oocytes were then cultured in OR3 medium for 72 h with fresh medium each day. Rates of L-lactate transport by wild-type or chimaeric MCTs were determined as described previously, either by following the change in intracellular pH using the ratiometric pH-sensitive fluorescent dye BCECF [2′-7′-bis(carboxyethyl)-5(6)-carboxyfluorescein] or by measuring uptake of [14C] substrate (L-lactate or pyruvate). For measurement of AR-C155858 sensitivity, ten oocytes were placed in a six-well plate containing 5 ml of uptake buffer [75 mM NaCl, 2 mM KCl, 0.82 mM MgCl2, 1 mM CaCl2 and 20 mM Mes (pH 6.0)] and allowed to pre-incubate for the required time (usually 45 min) with or without AR-C155858 as required. Five oocytes were removed and placed into 50 μl of uptake buffer containing L-[14C]lactate (0.5 mM, 7.4 MBq/ml) with or without AR-C155858 at the required concentration. Incubation at room temperature was continued for the period over which uptake was linear with time; this varied with the construct employed, as detailed in Supplementary Table S2. The oocytes were then rapidly washed five times with ice-cold uptake buffer and, after the final wash, each egg was transferred into a scintillation vial and homogenized in 100 μl of 2% (w/v) SDS by vigorous vortex-mixing. Scintillation fluid (10 ml of Emulsifier-Safe) was then added and [14C] assayed by scintillation counting.[2] For the determination of Km values for pyruvate and L-lactate, oocytes were equilibrated in incubation buffer [75 mM NaCl, 2 mM KCl, 0.82 mM MgCl2, 1 mM CaCl2 and 20 mM Tris/Hepes (pH 7.4)] for 5 min and then four oocytes incubated with 40 μl of uptake buffer containing [14C]-labelled L-lactate or pyruvate (7.4 MBq/ml) at a final concentration of 0.2, 0.5, 1, 2, 5, 20, 50 and 75 mM. Incubation was continued for the period over which uptake was linear with time (detailed in Supplementary Table S2), following which oocytes were washed and prepared for scintillation counting as above. Net MCT1-mediated uptake of L-lactate was determined by subtracting the uptake by water-injected oocytes determined at the same time.[2] MCT expression at the plasma membrane was confirmed by immunofluorescence microscopy of oocytes sectioned by embedding in chicken liver as described previously. |
| Cell Assay |
Measurement of MCT1 activity in erythrocytes [1]
L-Lactate transport into rat and rabbit erythrocytes was measured by monitoring the change in extracellular pH with a pH-sensitive electrode as described previously. The erythrocytes (5% haematocrit) were pre-incubated for 1 h at room temperature (22 °C) with or without AR-C155858 at the required concentration before assaying transport of L-lactate (10 mM) at 6 °C. Initial rates of transport were calculated by first-order regression analysis of the time course of pH change and converted into nmol of H+/min by determining the pH change induced by small additions of standardized NaOH. Measurement of MCT transport activity in Xenopus oocytes [1] cRNA was prepared and injected into X. laevis oocytes as described previously. For all assays, 10 ng of MCT cRNA ±10 ng of rat embigin cRNA in 9.2 nl of water were injected, with controls receiving just water. MCT and embigin expression at the plasma membrane of oocytes was confirmed by immunofluorescence microscopy as described previously. MCT kinetic assays were performed by monitoring intracellular pH with H+-sensitive dye BCECF [2′,7′-bis-(2-carboxyethyl)-5(6)-carboxyfluorescein] or by determining the uptake of L-[14C]lactate (7.4 MBq/ml). The uptake buffer contained 75 mM NaCl, 2 mM KCl, 0.82 mM MgCl2, 1 mM CaCl2 and 20 mM Tris/Hepes (pH 7.4). AR-C155858 inhibitor titrations were performed at pH 6 with oocytes pre-incubated for 45 min in a different uptake buffer (75 mM NaCl, 2 mM KCl, 0.82 mM MgCl2, 1 mM CaCl2 and 20 mM Mes, pH 6) containing the required concentration of AR-C155858 prior to measuring the uptake of L-[14C]lactate (0.5 mM) as described previously [38]. Unless stated otherwise, uptake was determined over 2.5 min for all MCT constructs except for MCT2trn with or without embigin and MCT2/1 with or without embigin, where 5 and 10 min were used respectively. We determined that these conditions represented the longest period over which uptake was linear with time (results not shown). |
| Animal Protocol |
Toxicokinetic/Toxicodynamic studies [3]
Effect of AR-C155858 on toxicokinetics and respiratory depression of intravenous γ-hydroxybutyric acid (GHB) [3] The effect of AR-C155858 on GHB-induced respiratory depression was studied using whole-body plethysmography similar to our previously published studies. Animals were allowed to acclimate to the plethysmography chambers for 45 minutes followed by collection of five baseline measurements of respiratory parameters over 15 minutes. GHB was administered intravenously as 200, 600 or 1500 mg/kg bolus with or without AR-C155858 (1 or 5 mg/kg i.v. bolus). In the GHB 600 mg/kg group, a lower dose of AR-C155858 (0.1 mg/kg i.v. bolus) was also administered. In all the treatment groups, AR-C155858 was administered 5 minutes after GHB administration. This experiment was performed at a similar time and in a similar manner to our previous study assessing respiratory effects of GHB alone; therefore, data from rats administered GHB 200, 600, and 1500 mg/kg alone were used from the previous publication for comparison purposes. The time of GHB administration was considered as time 0. Blood and urine samples were collected at intervals up to 8 hours after GHB administration. The respiratory parameters, breathing frequency, tidal volume, and minute volume (breathing frequency x tidal volume) were recorded at 2.5, 5, 7.5, 10, 15, 20, 25, and 30 minutes and every 15 minutes thereafter until 8 hours. In all groups of animals, GHB was administered as a 300 mg/ml solution in sterile water via the jugular vein cannula. The AR-C155858 bolus was administered as a 0.1, 1 or 2.5 mg/ml solution in 10 % cyclodextrin in normal saline via the jugular vein cannula. All the treatment groups included 3–6 animals and were compared with their respective GHB alone group to determine the effects of AR-C155858 on GHB-induced respiratory depression. A separate group of animals received AR-C155858 alone (1 mg/kg i.v. bolus) to study the effect of this inhibitor itself on respiration. Effect of AR-C155858 on GHB blood-brain partitioning at steady state [3] To assess the effect of AR-C155858 on the transport of GHB into the brain, GHB (400 mg/kg i.v. bolus followed by 208 mg/kg/hr i.v. infusion) was administered alone or in combination with AR-C155858 (5 mg/kg i.v. bolus) (n=4). The GHB dose was selected to produce steady-state GHB plasma concentrations of 800 µg/ml, similar to the high concentrations of GHB observed in rats after 600 mg/kg GHB i.v. used in the toxicokinetic study above. In addition this GHB concentration is similar to those seen in clinical cases of GHB overdose (5). The animals were euthanized at 4 h post GHB administration under isoflurane anesthesia followed by collection of blood and brain samples at steady state. Brain samples were immediately frozen in liquid nitrogen and stored at −80°C until analysis. Effect of AR-C155858 on oral GHB toxicokinetics Because GHB is commonly abused orally, the effect AR-C155858 was assessed on GHB toxicokinetics after oral administration in rats. Animals were administered GHB by oral gavage with or without AR-C155858 (5 mg/kg i.v. bolus). AR-C155858 was either administered 5 minutes or 1 hour post GHB administration (n = 4–6). In another group of animals, both AR-C155858 (10 mg/kg) and GHB (1500 mg/kg) were administered at the same time by oral gavage. Rats were fasted overnight before drug administration. Blood and urine samples were collected at intervals up to 15 hours after GHB administration. GHB alone was administered as a 300 mg/ml solution in water and AR-C155858 as a 2.5 mg/ml solution in 10% cyclodextrin in normal saline. |
| References |
|
| Additional Infomation |
In order to reconcile all of the results of the present study, we propose the scheme shown in Figure 7. The key features of this proposal are as follows.
(i) MCT2 binds preferentially to embigin over endogenous basigin, with the binding to the latter requiring an interaction between the C-termini of both proteins that is not required by MCT1. Thus MCT1trn, but not MCT2trn, expresses well in the absence of co-expressed embigin. (ii) MCT1 associates with embigin in the absence of basigin, but prefers the latter as binding partner when both are present, unless the C-terminal tail of MCT2 replaces the MCT1 C-terminus (MCT1/2c) when binding to embigin is promoted. (iii) When bound to embigin, the binding affinity of AR-C155858 to MCT2, but not MCT1, is greatly reduced and this effect is independent of the presence of the C-terminal tail of MCT2. Our data emphasize that the potency with which AR-C155858 inhibits MCT2 is dependent on the ancillary protein with which it associates. In mammalian cells, embigin is the preferred endogenous binding partner of MCT2 [22] and thus MCT2-mediated lactate transport is likely to be considerably less sensitive to inhibition by AR-C155858 than that mediated by MCT1. This may justify the cautious use of AR-C155858 to dissect out the different roles of MCT1 (very sensitive to AR-C155858), MCT2 (less sensitive to AR-C155858) and MCT4 (insensitive to AR-C155858) in the metabolism of tissues such as the brain, as has been reported by Bröer and colleagues [41]. However, the development of totally isoform-specific inhibitors of MCTs that are not influenced by the associated ancillary protein is clearly desirable. [1] Previously, a new class of specific and extremely potent inhibitors of MCT1 have been discovered by AstraZeneca that were reported to show no binding to MCT4 and exhibit lower-affinity binding to MCT2. The Kd values for these inhibitors binding to endogenous MCT1 in rat and human cells was found to be in the low nanomolar region or less, as was also found for MCT1 expressed in Ins-1 cells that contain little or no endogenous MCT1. Similar Kd values were determined for MCT1 expressed in yeast, and the Ki value of AR-C155858 for MCT1 we have determined in rat erythrocytes of 2.3±1.4 nM is entirely consistent with the Ki of 1.2 nM for binding of AR-C155858 to human erythrocyte MCT1 derived from radioligand-binding experiments. [2] In summary, the novel and highly potent inhibitor, AR-C155858 increases renal and total clearance of GHB following both intravenous and oral administration in rats. AR-C155858 also results in significant improvement of GHB-induced respiratory depression which may be mediated by inhibition of its renal reabsorption and brain uptake, both processes mediated by MCTs. Our studies demonstrate proof-of-concept in utilizing MCT inhibition as a potential treatment strategy by improving GHB-induced respiratory depression which leads to death in cases of GHB overdose. [3] |
| Molecular Formula |
C21H27N5O5S
|
|---|---|
| Molecular Weight |
461.5346
|
| Exact Mass |
461.173
|
| Elemental Analysis |
C, 54.65; H, 5.90; N, 15.17; O, 17.33; S, 6.95
|
| CAS # |
496791-37-8
|
| PubChem CID |
10226546
|
| Appearance |
Off-white to light yellow solid powder
|
| Density |
1.4±0.1 g/cm3
|
| Boiling Point |
763.1±70.0 °C at 760 mmHg
|
| Flash Point |
415.3±35.7 °C
|
| Vapour Pressure |
0.0±2.7 mmHg at 25°C
|
| Index of Refraction |
1.634
|
| LogP |
0.93
|
| Hydrogen Bond Donor Count |
2
|
| Hydrogen Bond Acceptor Count |
7
|
| Rotatable Bond Count |
5
|
| Heavy Atom Count |
32
|
| Complexity |
769
|
| Defined Atom Stereocenter Count |
1
|
| SMILES |
CC1=C(C(=NN1)C)CC2=C(C3=C(S2)N(C(=O)N(C3=O)C)CC(C)C)C(=O)N4C[C@@H](CO4)O
|
| InChi Key |
ISIVOJWVBJIOFM-ZDUSSCGKSA-N
|
| InChi Code |
InChI=1S/C21H27N5O5S/c1-10(2)7-25-20-17(18(28)24(5)21(25)30)16(19(29)26-8-13(27)9-31-26)15(32-20)6-14-11(3)22-23-12(14)4/h10,13,27H,6-9H2,1-5H3,(H,22,23)/t13-/m0/s1
|
| Chemical Name |
(S)-6-((3,5-dimethyl-1H-pyrazol-4-yl)methyl)-5-(4-hydroxyisoxazolidine-2-carbonyl)-1-isobutyl-3-methylthieno[2,3-d]pyrimidine-2,4(1H,3H)-dione
|
| Synonyms |
ARC155858; AR-C155858; 496791-37-8; (S)-6-((3,5-dimethyl-1H-pyrazol-4-yl)methyl)-5-(4-hydroxyisoxazolidine-2-carbonyl)-1-isobutyl-3-methylthieno[2,3-d]pyrimidine-2,4(1H,3H)-dione; (S)-6-[(3,5-Dimethyl-1H-pyrazol-4-yl)methyl]-5-[(4-hydroxyisoxazolidin-2-yl)carbonyl]-1-isobutyl-3-methylthieno[2,3-d]pyrimidine-2,4(1H,3H)-dione; 6-[(3,5-dimethyl-1H-pyrazol-4-yl)methyl]-5-[(4S)-4-hydroxy-1,2-oxazolidine-2-carbonyl]-3-methyl-1-(2-methylpropyl)thieno[2,3-d]pyrimidine-2,4-dione; AR-C 155858; 6-[(3,5-dimethyl-1H-pyrazol-4-yl)methyl]-5-[(4S)-4-hydroxy-1,2-oxazolidine-2-carbonyl]-3-methyl-1-(2-methylpropyl)-1H,2H,3H,4H-thieno[2,3-d]pyrimidine-2,4-dione; ARC 155858.
|
| HS Tariff Code |
2934.99.9001
|
| Storage |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| Shipping Condition |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| Solubility (In Vitro) |
DMSO : ~70 mg/mL (~151.67 mM)
|
|---|---|
| Solubility (In Vivo) |
Solubility in Formulation 1: ≥ 2.75 mg/mL (5.96 mM) (saturation unknown) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (add these co-solvents sequentially from left to right, and one by one), clear solution.
For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 27.5 mg/mL clear DMSO stock solution to 400 μL PEG300 and mix evenly; then add 50 μL Tween-80 to the above solution and mix evenly; then add 450 μL normal saline to adjust the volume to 1 mL. Preparation of saline: Dissolve 0.9 g of sodium chloride in 100 mL ddH₂ O to obtain a clear solution. Solubility in Formulation 2: ≥ 2.75 mg/mL (5.96 mM) (saturation unknown) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (add these co-solvents sequentially from left to right, and one by one), clear solution. For example, if 1 mL of working solution is to be prepared, you can add 100 μL of 27.5 mg/mL clear DMSO stock solution to 900 μL of 20% SBE-β-CD physiological saline solution and mix evenly. Preparation of 20% SBE-β-CD in Saline (4°C,1 week): Dissolve 2 g SBE-β-CD in 10 mL saline to obtain a clear solution. (Please use freshly prepared in vivo formulations for optimal results.) |
| Preparing Stock Solutions | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.1667 mL | 10.8335 mL | 21.6671 mL | |
| 5 mM | 0.4333 mL | 2.1667 mL | 4.3334 mL | |
| 10 mM | 0.2167 mL | 1.0834 mL | 2.1667 mL |
*Note: Please select an appropriate solvent for the preparation of stock solution based on your experiment needs. For most products, DMSO can be used for preparing stock solutions (e.g. 5 mM, 10 mM, or 20 mM concentration); some products with high aqueous solubility may be dissolved in water directly. Solubility information is available at the above Solubility Data section. Once the stock solution is prepared, aliquot it to routine usage volumes and store at -20°C or -80°C. Avoid repeated freeze and thaw cycles.
Calculation results
Working concentration: mg/mL;
Method for preparing DMSO stock solution: mg drug pre-dissolved in μL DMSO (stock solution concentration mg/mL). Please contact us first if the concentration exceeds the DMSO solubility of the batch of drug.
Method for preparing in vivo formulation::Take μL DMSO stock solution, next add μL PEG300, mix and clarify, next addμL Tween 80, mix and clarify, next add μL ddH2O,mix and clarify.
(1) Please be sure that the solution is clear before the addition of next solvent. Dissolution methods like vortex, ultrasound or warming and heat may be used to aid dissolving.
(2) Be sure to add the solvent(s) in order.
|
|
|
Products are for research use only; We do not sell to patients
Copyright 2020 InvivoChem LLC | All Rights Reserved



























 COA
COA